 |
 |
|
|
| |
 |
植物資源の生産と利用 |
|
| 1. | 食糧および産業に関する植物遺伝資源 |
| 1.1. | 農業の生産性と持続性の問題点 |
| 1.1.1. | 食糧危機 |
| 1.1.2. | 食糧の量と質:栄養不足と食品安全性 |
| 1.1.3. | 環境破壊と天然資源の枯渇 |
| 1.1.4. | 緑の革命 |
| 1.1.5. | 品種と遺伝資源 |
| 1.2. | 植物遺伝資源と世界 |
| 1.2.1. | 遺伝資源の重要性 |
| 1.2.2. | 遺伝資源、バイオテクノロジーと世界 |
| 1.2.3. | 遺伝資源の保全と植物科学 |
| 1.2.4. | 遺伝資源の利用とバイオテクノロジー |
| 1.2.5. | 最後に |
| 2. | 森林資源 |
| 2.1. | はじめに |
| 2.2. | 世界における木材、紙パルプの生産量と消費 |
| 2.3. | 紙パルプ原料の品種開発(育種) |
| 2.3.1. | 長・短期の育種戦略 |
| 2.3.2. | 第1段階:量の育種 |
| 2.3.3. | 第2段階:質の育種 |
| 2.4. | まとめ |
| 3. | 工業用資源 |
| 3.1. | デンプン |
| 3.1.1. | はじめに |
| 3.1.2. | デンプンの生産量と種類および一般的性質 |
| 3.1.2.1. | デンプンの生産量 |
| 3.1.2.2. | デンプンの種類と一般的性質 |
| 3.1.3. | デンプンの生産と加工技術および利用 |
| 3.1.3.1. | デンプンの生産 |
| 3.1.3.2. | デンプンの加工技術と利用 |
| 3.1.4. | 今後の課題 |
| 3.1.4.1. | 新しいデンプンの作出 |
| 3.1.4.2. | デンプンの生分解性プラスチック分野への利用ほか |
| 3.2. | リグノセルロース |
| 3.2.1. | 資源とその利用の概要 |
| 3.2.2. | リグノセルロースの変換と利用 |
| 3.2.2.1. | 全体利用 |
| 3.2.2.2. | 成分利用 |
| 3.2.3. | おわりに |
| 3.3. | 油脂 |
| 3.3.1. | はじめに |
| 3.3.2. | 生産量および原料生産国 |
| 3.3.3. | 需要動向 |
| 3.3.4. | 採油および精製技術 |
| 3.3.5. | 食用油脂加工技術 |
| 3.3.5.1. | 水素添加(硬化) |
| 3.3.5.2. | エステル交換 |
| 3.3.5.3. | 分別 |
| 3.3.6. | 工業分野への利用 |
| 3.3.6.1. | 脂肪酸製造技術 |
| 3.3.6.2. | 高級アルコール製造技術 |
| 3.3.6.3. | メチルエステル製造技術 |
| 3.3.6.4. | 脂肪酸誘導体とその応用 |
| 3.3.6.5. | 高級アルコール誘導体とその応用 |
| 3.3.6.6. | 油脂の直接利用 |
| 3.3.7. | おわりに |
| 3.4. | 天然ゴム |
| 3.4.1. | 天然ゴムの種類 |
| 3.4.2. | 天然ゴムの構造と非ゴム成分 |
| 3.4.3. | 天然ゴムの精製 |
| 3.4.4. | 天然ゴムの非ゴム成分と物性 |
| 3.4.5. | 天然ゴムの改質 |
| 3.4.6. | 高純度天然ゴムの改質 |
| 3.4.7. | ゴムの樹の品種改良 |
| 3.5. | 繊維 |
| 3.5.1. | 植物および植物由来繊維の種類 |
| 3.5.1.1. | 植物繊維 |
|
|
| 3.5.1.2. | 植物由来繊維 |
| 3.5.2. | 植物および植物由来繊維の生産量 |
| 3.5.2.1. | 植物繊維 |
| 3.5.2.2. | 植物由来繊維 |
| 3.5.3. | 植物および植物由来繊維の用途 |
| 3.5.3.1. | 植物繊維 |
| 3.5.3.2. | 植物由来繊維 |
| 3.5.4. | 植物および植物由来繊維の遺伝子操作 |
| 3.5.4.1. | 植物繊維 |
| 3.5.4.2. | 植物由来繊維 |
| 3.6. | 色素、香料、生薬、タンパク質 |
| 3.6.1. | はじめに |
| 3.6.2. | 色素 |
| 3.6.2.1. | 植物色素の種類と原料植物 |
| 3.6.2.2. | 植物系食品色素の生産動向 |
| 3.6.2.3. | 植物染料の動向 |
| 3.6.2.4. | 植物色素の課題 |
| 3.6.3. | 香料 |
| 3.6.3.1. | 植物香料の種類と用途 |
| 3.6.3.2. | フレーバーの動向 |
| 3.6.3.3. | フレグランスの動向 |
| 3.6.3.4. | 植物香料の課題 |
| 3.6.4. | 生薬 |
| 3.6.4.1. | 植物生薬の種類と生産量 |
| 3.6.4.2. | 植物生薬の課題 |
| 3.6.5. | タンパク質 |
| 3.6.5.1. | 植物性タンパク質の種類と生産量 |
| 3.6.5.2. | 遺伝子組換え植物による動物性タンパク質の生産 |
| 3.6.6. | おわりに |
| 4. | 未利用バイオマス |
| 4.1. | はじめに |
| 4.2. | バイオマスの種類と用途 |
| 4.2.1. | バイオマスの種類 |
| 4.2.2. | バイオマスの用途 |
| 4.2.3. | バイオマスの潜在的存在量 |
| 4.3. | 未利用バイオマスの発生量と利用の現状 |
| 4.3.1. | 未利用バイオマスの利用と問題点 |
| 4.3.2. | 廃棄系バイオマスの発生量と再利用の現状 |
| 4.3.3. | 植物系未利用バイオマスの発生量と再利用の現状 |
| 4.4. | 未利用バイオマスの利用拡大の課題 |
| 5. | 植生環境 |
| 5.1. | 植生の変遷 |
| 5.1.1. | はじめに |
| 5.1.2. | 植生のタイプと純一次生産量 |
| 5.1.3. | 熱帯林減少 |
| 5.1.3.1. | 熱帯林減少の現状 |
| 5.1.3.2. | 熱帯林減少の原因 |
| 5.1.3.3. | 熱帯林減少の影響 |
| 5.1.4. | 酸性雨 |
| 5.1.4.1. | 酸性雨生成過程 |
| 5.1.4.2. | 酸性雨の影響 |
| 5.1.5. | 砂漠化 |
| 5.1.5.1. | 砂漠化の定義 |
| 5.1.5.2. | 砂漠化の現状 |
| 5.1.5.3. | 砂漠化の原因 |
| 5.1.6. | 耕地拡張 |
| 5.1.7. | 地球温暖化の影響 |
| 5.1.8. | 植物バイオ育種への提言 |
| 5.2. | 土壌劣化 |
| 5.2.1. | はじめに |
| 5.2.2. | 砂漠化、塩類集積、草原化 |
| 5.2.3. | 表土流亡、土壌浸食 |
| 5.2.4. | 土壌への有害物質の蓄積 |
| 5.2.5. | 地力の低下 |
| 5.2.6. | 酸性雨 |
|
| |
 |
植物代謝工学のための分子改良技術 |
|
| 1. | ゲノム情報工学 |
| 1.1. | ゲノムプロジェクト |
| 1.1.1. | はじめに |
| 1.1.2. | 植物のゲノム解析 |
| 1.1.2.1. | モデル生物のゲノム解読 |
| 1.1.2.2. | ゲノム解読の方法論(原核生物の場合) |
| 1.1.2.3. | ゲノム解読の方法論(真核生物の場合) |
| 1.1.2.4. | 遺伝子構造予測 |
| 1.1.2.5. | ラン藻のゲノム解読 |
| 1.1.2.6. | シロイヌナズナのゲノム解読 |
| 1.1.2.7. | イネのゲノム解読 |
| 1.1.2.8. | マメ科植物および根粒菌のゲノム解読 |
| 1.1.2.9. | ESTの整備 |
| 1.1.3. | ゲノム情報から得られる有用知見 |
| 1.1.4. | ゲノム情報利用の展望 |
| 1.2. | トランスクリプトーム解析とDNAマイクロアレイ |
| 1.2.1. | はじめに |
| 1.2.2. | トランスクリプトーム解析の現状 |
| 1.2.2.1. | cDNAのランダムシーケンス法による頻度情報の記録 |
| 1.2.2.2. | ディスプレイ法 |
| 1.2.2.3. | ディファレンシャルハイブリダイゼーション |
| 1.2.2.4. | 定量的PCR |
| 1.2.3. | DNAマイクロアレイ技術の実際 |
| 1.2.3.1. | マイクロアレイ用DNA(プローブDNA)の調製 |
| 1.2.3.2. | RNAサンプルからのターゲットDNAの調製 |
| 1.2.3.3. | マイクロアレイの作製とハイブリダイゼーション |
| 1.2.3.4. | シグナル検出とスポットの定量 |
| 1.2.3.5. | データ解析 |
| 1.2.4. | 植物研究分野におけるバイオインフォマティクスの利用とマイクロアレイ研究の展望 |
| 1.2.4.1. | 植物分野におけるバイオインフォマティクス関連のWebサイト |
| 1.2.4.2. | 植物研究分野におけるマイクロアレイ研究の展望 |
| 1.3. | 遺伝子マッピング |
| 1.3.1. | 遺伝子のフィジカルマッピングとその意義 |
| 1.3.1.1. | フィジカルマッピングの意義 |
| 1.3.1.2. | FISH法 |
| 1.3.2. | マッピングに必要な染色体地図作製法と各種染色体地図 |
| 1.3.2.1. | 画像解析法 |
| 1.3.2.2. | 各種染色体地図 |
| 1.3.3. | 各種FISH法 |
| 1.3.3.1. | 多重遺伝子族のマッピング |
| 1.3.3.2. | 種特異的反復配列のマッピング |
| 1.3.3.3. | 単一コピー遺伝子のマッピング |
| 1.3.3.4. | マルチカラーFISH (M-FISH)法 |
| 1.3.3.5. | EDF-FISH法 |
| 1.3.3.6. | RCA法による単一塩基対変異の検出 |
| 1.3.4. | マッピング技術の応用例 |
| 1.3.4.1. | rDNA多型の検出 |
| 1.3.4.2. | 反復配列の変異の検出 |
| 1.3.4.3. | 遺伝子の位置関係の検出 |
| 1.3.4.4. | 形質転換体における外来遺伝子の検出 |
| 2. | 遺伝子導入技術 |
| 2.1. | 核染色体への外来遺伝子の導入法 |
| 2.1.1. | ベクターの種類とマーカー遺伝子 |
| 2.1.1.1. | はじめに |
| 2.1.1.2. | 遺伝子導入法 |
| 2.1.1.2.1. | 直接法 |
| 2.1.1.2.2. | 直接導入法に用いられるベクター |
| 2.1.1.3. | アグロバクテリウムを媒介として用いる方法 |
| 2.1.1.4. | アグロバクテリウムの感染機構 |
| 2.1.1.5. | アグロバクテリウムのTiプラスミド由来ベクター |
| 2.1.1.5.1. | 中間ベクター |
| 2.1.1.5.2. | バイナリーベクター |
| 2.1.1.5.3. | 選択マーカー遺伝子が除去されるベクター |
| 2.1.1.6. | アグロバクテリウムの宿主範囲 |
| 2.1.1.7. | 選択マーカー遺伝子 |
| 2.1.1.7.1. | 抗生物質または除草剤耐性遺伝子 |
| 2.1.1.7.2. | 薬剤耐性以外のマーカー遺伝子 |
| 2.1.1.8. | 問題点と今後の展望 |
| 2.1.2. | 個体再生技術と代謝工学の対象となる植物 |
| 2.1.2.1. | 個体再生技術 |
| 2.1.2.1.1. | 不定芽形成(不定芽誘導法) |
| 2.1.2.1.2. | 苗条原基法 |
| 2.1.2.1.3. | 不定胚形成(embryogenesis)(不定胚誘導法) |
| 2.1.2.1.4. | 遺伝子導入による再分化技術 |
| 2.1.2.1.5. | 個体再生技術の形質転換への利用 |
| 2.1.2.1.6. | 再生技術を用いない形質転換法 |
| 2.1.2.1.7. | 再生技術の形質転換への利用 |
| 2.1.2.2. | 代謝工学の対象となる植物 |
| 2.1.2.2.1. | ユーカリ |
| 2.1.2.2.2. | ダイズ |
| 2.1.2.2.3. | サツマイモ |
| 2.1.2.2.4. | イネ |
| 2.1.2.2.5. | ゴムの木 |
| 2.1.2.2.6. | ナタネ |
| 2.1.2.3. | 代謝工学の対象となる代謝経路 |
| 2.1.2.4. | 今後の展望 |
| 2.2. | 核外小器官への遺伝子導入法 |
| 2.2.1. | オルガネラ形質転換とは |
| 2.2.2. | 葉緑体形質転換 |
| 2.2.3. | 葉緑体逆遺伝学 |
| 2.2.4. | 光合成機能改変 |
| 2.2.5. | 母性遺伝 |
| 2.2.6. | 葉緑体工場 |
| 2.2.7. | 葉緑体形質転換の現状 |
|
|
| 2.2.8. | ゲノム安定性に関する課題 |
| 2.2.9. | ミトコンドリアゲノムの形質転換 |
| 2.3. | 多重遺伝子導入技術 |
| 2.3.1. | はじめに |
| 2.3.2. | 交配法による複数遺伝子の導入 |
| 2.3.3. | 逐次形質転換法 |
| 2.3.4. | 複数ベクターによる共形質転換法 |
| 2.3.5. | 単一ベクターによる多重遺伝子導入法 |
| 2.3.5.1. | 長鎖DNAを導入する技術 |
| 2.3.5.2. | 複数のDNA断片を効率よくベクターに連結する技術の開発 |
| 2.3.6. | おわりに |
| 3. | 遺伝子発現制御技術 |
| 3.1. | 核遺伝子 |
| 3.1.1. | 有用プロモーター |
| 3.1.1.1. | プロモーターの基本配列 |
| 3.1.1.2. | 5’上流シス配列・エンハンサー配列 |
| 3.1.1.3. | 有用プロモーターの現状 |
| 3.1.1.3.1. | 構成的高発現プロモーター |
| 3.1.1.3.2. | 組織特異的プロモーター |
| 3.1.1.3.3. | 誘導発現プロモーター |
| 3.1.1.4. | 有用プロモーターの単離法 |
| 3.1.1.5. | 外来遺伝子発現のポジション効果を抑制する技術 |
| 3.1.1.5.1. | 染色体DNAの高次構造が遺伝子発現に及ぼす影響 |
| 3.1.1.5.2. | インスレーターの利用 |
| 3.1.1.5.3. | MARの利用 |
| 3.1.1.6. | 外来遺伝子発現のサイレンシング |
| 3.1.1.7. | おわりに |
| 3.1.2. | 翻訳効率 |
| 3.1.2.1. | はじめに |
| 3.1.2.2. | 植物における翻訳機構 |
| 3.1.2.3. | 遺伝子の発現を翻訳レベルで高めるアプローチ |
| 3.1.2.3.1. | 植物ウイルス由来の5’UTR(5’リーダー配列) |
| 3.1.2.3.2. | 植物の遺伝子由来の5’UTR |
| 3.1.2.4. | 次世代の植物代謝工学につながる技術 |
| 3.1.2.4.1. | mRNAの安定化 |
| 3.1.2.4.2. | ポリシストロン(IRES配列) |
| 3.1.2.4.3. | ストレス時の翻訳抑制の回避 |
| 3.1.2.4. | 今後の展望 |
| 3.2. | 貯蔵器官および細胞内小器官(葉緑体)特異的な発現 |
| 3.2.1. | はじめに |
| 3.2.2. | バイオリアクターとしての種子 |
| 3.2.3. | 種子での異種タンパク質蓄積の試み |
| 3.2.4. | 種子での異種タンパク質高蓄積の試み |
| 3.2.5. | バイオリアクターとしての葉緑体 |
| 3.2.6. | 葉緑体での異種タンパク質蓄積の試み |
| 3.2.6.1. | 微生物由来の遺伝子・オペロン |
| 3.2.6.2. | 動物由来の遺伝子・ジスルフィド結合 |
| 3.2.7. | 葉緑体での工業原料生産の試み |
| 3.2.8. | 次世代の葉緑体工学につながる技術 |
| 3.3. | 遺伝子発現抑制技術 |
| 3.3.1. | はじめに |
| 3.3.2. | 変異株の単離 |
| 3.3.3. | 相同配列組換え、トランスポゾンあるいはT-DNAを用いた遺伝子破壊 |
| 3.3.4. | アンチセンス法 |
| 3.3.5. | コサプレッションとRNAi |
| 3.3.5.1. | dsRNA発現ベクターの構築 |
| 3.3.5.2. | RNAiと標的の配列 |
| 3.3.5.3. | RNAiの効率 |
| 3.3.5.4. | RNAiの検出 |
| 3.3.5.5. | TGS |
| 3.3.6. | 最後に |
| 3.4. | 植物糖鎖工学 |
| 3.4.1. | はじめに |
| 3.4.1.1. | 植物N―結合型糖鎖 |
| 3.4.1.2. | 植物O―結合型糖鎖 |
| 3.4.2. | 植物における糖鎖プロセシング |
| 3.4.3. | 植物の分子糖鎖生物学 |
| 3.4.4. | 植物糖鎖の生理学的役割 |
| 3.4.5. | 植物糖鎖とアレルギー |
| 3.4.6. | 植物糖鎖構造のヒト型化 |
| 4. | 植物のポストゲノムの動向 |
| 4.1. | 植物のプロテオーム解析 |
| 4.1.1. | プロテオーム解析とは? |
| 4.1.2. | プロテオーム解析の原理 |
| 4.1.3. | 植物におけるプロテオーム解析 |
| 4.1.3.1. | 葉緑体のプロテオミクス |
| 4.1.3.2. | 植物の分化 |
| 4.1.3.3. | 環境応答 |
| 4.1.4. | プロテオーム解析の課題と展望 |
| 4.2. | 植物のメタボローム解析 |
| 4.2.1. | 植物メタボローム解析とは |
| 4.2.2. | メタボローム解析のための分析戦略 |
| 4.2.2.1. | メタボローム解析のための分析に求められる基準 |
| 4.2.2.2. | HPLCおよびGCの一般的特徴 |
| 4.2.2.3. | 質量分析(MS)の一般的特徴 |
| 4.2.2.4. | 代謝産物分析の実際 |
| 4.2.3. | 今後の展望 |
| 5. | 植物の分子改良技術基本特許の状況 |
| 5.1. | 植物の分子改良技術 |
| 5.1.1. | 遺伝子導入技術の現状と課題 |
| 5.1.2. | 外来遺伝子の発現調節技術の現状と課題 |
| 5.1.3. | 遺伝子導入細胞の選抜技術の現状と課題 |
| 5.2. | 植物の分子改良技術の基本特許 |
|
| |
 |
一次生産向上のための代謝工学 |
|
| 1. | 光エネルギー変換機構 |
| 1.1. | 葉緑体構造 |
| 1.2. | 光エネルギー捕捉 |
| 1.2.1. | 光を捕獲するアンテナ |
| 1.2.2. | クロロフィルの光励起 |
| 1.2.3. | クロロフィル蛍光 |
| 1.2.4. | 光化学系反応中心における反応 |
| 1.3. | 光エネルギーから化学エネルギーへの変換を担う成分 |
| 1.3.1. | 光化学系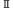 |
| 1.3.2. | プラストキノン |
| 1.3.3. | シトクロムb6/f複合体 |
| 1.3.4. | プラストシアニン |
| 1.3.5. | 光化学系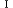 |
| 1.3.6. | フェレドキシン |
| 1.3.7. | フェレドキシン-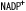 オキシドレダクターゼ(FNR) オキシドレダクターゼ(FNR) |
| 1.3.8. | ATPシンターゼ |
| 1.4. | 光合成電子伝達反応(非循環的電子伝達反応) |
| 1.5. | ATP合成 |
| 1.6. | チラコイド膜におけるその他の電子伝達反応 |
| 1.6.1. | 光合成に必要な電子/プロトンの化学量論比 |
| 1.6.2. | 光化学系Iにおける循環的電子伝達反応 |
| 1.6.3. | Qサイクル |
| 1.6.4. | PSIにおける酸素への電子伝達反応(water-water サイクル) |
| 1.7. | 光エネルギー変換系における代謝制御 |
| 1.7.1. | 光化学系IIのダウンレギュレーション |
| 1.7.2. | キサントフィルサイクル |
| 1.7.3. | 過剰光エネルギー散逸系としてのwater-water サイクル |
| 1.7.4. | シトクロム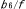 複合体による電子伝達制御 複合体による電子伝達制御 |
| 1.7.5. | チオレドキシンによるATPシンターゼの活性制御 |
| 1.7.6. | 生体内の酸化剤によるATPシンターゼの活性調節 |
| 1.8. | おわりに |
| 2. | 炭素代謝 |
| 2.1. | はじめに |
| 2.2. | 光合成における炭素固定・還元系 |
| 2.3. | 生産性におけるRuBisCOの役割 |
| 2.3.1. | PCRサイクル酵素の一般的特徴 |
| 2.3.2. | RuBisCO |
| 2.4. | 光呼吸 |
| 2.4.1. | 光呼吸代謝系 |
| 2.4.2. | 光呼吸の生理 |
| 2.5. | 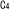 -経路: -経路: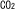 同化のための補助経路 同化のための補助経路 |
| 2.6. | 光合成の律速因子と律速解除による生産性向上のためのバイオテクノロジー |
| 3. | 窒素代謝 |
| 3.1. | はじめに |
| 3.2. | 硝酸イオンの吸収と輸送 |
| 3.3. | 硝酸還元 |
| 3.4. | アンモニウムイオンの吸収 |
|
|
| 3.5. | アンモニア同化 |
| 3.6. | 硝酸イオンによる遺伝子制御ネットワーク |
| 3.7. | 窒素栄養と形態的応答 |
| 3.8. | アミノ酸代謝 |
| 3.9. |  植物と植物 植物と植物 |
| 3.10. | 窒素化合物の輸送・転流 |
| 3.11. | 窒素栄養による遺伝子発現制御 |
| 3.12. | 翻訳後の制御機構 |
| 3.13. | 代謝工学面への応用 |
| 4. | ソース・シンク相互作用 |
| 4.1. | はじめに |
| 4.2. | ソースとシンクの多様性 |
| 4.3. | ソースでの炭素代謝 |
| 4.3.1. | デンプン合成 |
| 4.3.2. | ショ糖の合成 |
| 4.3.3. | デンプンの分解 |
| 4.3.4. | ショ糖・デンプン代謝関連の変異株・アンチセンス形質転換植物を用いた研究 |
| 4.4. | 師管への糖の供給(ローディング) |
| 4.4.1. | アポプラスティックな経路 |
| 4.4.2. | シンプラスティックな輸送 |
| 4.5. | 師管内の糖の輸送 |
| 4.6. | シンク力 |
| 4.7. | シンクによる師管からの糖の取り出し(アンローディング) |
| 4.7.1. | マメや穀類の種子 |
| 4.7.2. | サトウダイコン |
| 4.7.3. | トマトの果実 |
| 4.7.4. | ジャガイモ |
| 4.7.5. | トウモロコシやオオムギの胚乳のデンプン合成 |
| 4.7.6. | シンクが多数ある場合 |
| 4.7.7. | シンクからソースに変わる葉の場合 |
| 4.7.8. | ショ糖の分解 |
| 4.8. | 糖を介したソース・シンク間の情報伝達 |
| 4.9. | 植物の生産性についてのソース・シンクからの展望 |
| 5. | 微量元素(イオウを含めて)の吸収と代謝 |
| 5.1. | はじめに |
| 5.2. | イオウ |
| 5.2.1. | 植物および環境中のイオウ化合物 |
| 5.2.2. | 硫酸イオンの吸収と輸送 |
| 5.2.3. | 硫酸イオンの還元同化とシステイン合成 |
| 5.2.4. | メチオニン生合成 |
| 5.2.5. | グルタチオン生合成 |
| 5.2.6. | そのほかのイオウ化合物 |
| 5.2.7. | イオウ栄養と生育 |
| 5.3. | 鉄 |
| 5.3.1. | 土壌中の鉄化合物 |
| 5.3.2. | 鉄の吸収 |
| 5.3.3. | 鉄の役割 |
| 5.4. | そのほかの微量元素 |
|
| |
 |
ストレス応答機構の分子改良 |
|
| 1. | 塩・乾燥ストレス耐性植物 |
| 1.1. | 植物が受ける塩・乾燥ストレス |
| 1.2. | 植物の塩・乾燥ストレスに対する適応機構 |
| 1.3. | 塩・乾燥ストレス耐性植物作製のための標的分子 |
| 1.4. | 浸透圧ストレス耐性を賦与する機能分子の改良 |
| 1.5. | 光酸素ストレス耐性を賦与する機能分子の改良 |
| 1.6. | イオンストレスに対する耐性を賦与する機能分子の改良 |
| 1.7. | 塩・乾燥ストレスの適応応答に関与するシグナル伝達分子の改良 |
| 1.8. | 塩・乾燥ストレス耐性に関与する機能未知のタンパク質分子の改良 |
| 1.9. | 塩・乾燥ストレス耐性植物作製のための今後の戦略 |
| 2. | 温度ストレスに対する応答と温度ストレス耐性植物 |
| 2.1. | はじめに |
| 2.2. | 低温ストレスに対する生理応答 |
| 2.3. | 低温ストレスに対する遺伝子レベルでの応答 |
| 2.4. | 低温ストレス応答における情報伝達系 |
| 2.5. | 耐冷性・耐凍性植物の作出 |
| 2.6. | 高温ストレスに対する生理応答 |
| 2.6.1. | HSP100 |
| 2.6.2. | HSP90 |
| 2.6.3. | HSP70 |
| 2.6.4. | HSP60 |
| 2.6.5. | HSP20 |
| 2.7. | 高温ストレスに対する遺伝子応答とその制御 |
| 2.8. | 高温耐性植物の作出 |
| 3. | 光・酸素毒耐性植物 |
| 3.1. | はじめに |
| 3.2. | AOSの生成 |
| 3.2.1. | おもなオルガネラでのAOS生成系 |
| 3.2.2. | 種々の環境要因のAOS生成に及ぼす影響 |
| 3.3. | AOSの消去系 |
| 3.3.1. | 抗酸化物質 |
| 3.3.2. | 抗酸化酵素 |
| 3.3.3. | 藻類のAOS消去および耐性機構 |
| 3.3.4. | AOS消去機構のストレス応答性 |
| 3.3.5. | AOS消去機構の問題点 |
| 3.4. | 光・酸素毒耐性の強化を目指した形質転換植物 |
| 3.4.1. | AOS消去酵素の制御 |
| 3.4.2. | 抗酸化物質の制御 |
| 3.4.3. | その他の代謝系の制御 |
| 3.5. | 遺伝子ターゲティングのストラテジー ‘今後の展望’ |
| 3.5.1. | 新たなストレス耐性関連遺伝子の探索と導入 |
| 3.5.2. | オルガネラ内での局在化(ミクロターゲティング) |
| 3.5.3. | 導入遺伝子の発現制御 |
| 3.5.4. | 転写後調節機構の改変 |
| 3.5.5. | 遺伝子発現のシグナル伝達系の改変 |
|
|
| 3.5.6. | 多重遺伝子導入 |
| 3.5.7. | 耐性植物の評価法の改良 |
| 3.6. | おわりに |
| 4. | 病害虫耐性植物の分子育種 |
| 4.1. | はじめに |
| 4.2. | 植物の病害抵抗性について |
| 4.2.1. | 過敏感反応 |
| 4.2.2. | 誘導抵抗性 |
| 4.2.3. | 抵抗性遺伝子 |
| 4.2.4. | PRタンパク質 |
| 4.2.5. | ファイトアレキシン |
| 4.3. | これまでに導入されてきた遺伝子 |
| 4.3.1. | 抵抗性遺伝子 |
| 4.3.2. | PRタンパク質遺伝子 |
| 4.3.3. | リボソーム不活化タンパク質遺伝子 |
| 4.3.4. | 病原体由来の遺伝子 |
| 4.3.5. | 抗菌性ペプチド |
| 4.4. | 新規なアプローチによる形質転換植物 |
| 4.4.1. |  遺伝子 遺伝子 |
| 4.4.2. | カルモジュリン遺伝子 |
| 4.4.3. | 病原体誘導型プロモーター:エリシター遺伝子 |
| 4.5. | 耐虫性について |
| 4.5.1. | Bt遺伝子 |
| 4.5.2. | プロテアーゼインヒビター |
| 4.5.3. | 食害により放出される揮発性シグナル |
| 4.6. | これからの病害抵抗性植物の作出 |
| 4.7. | おわりに |
| 5. | 農薬等負荷軽減植物 |
| 5.1. | はじめに |
| 5.2. | 除草剤耐性作物 |
| 5.3. | 環境モニタリングおよび汚染軽減植物 |
| 5.3.1. | ダイオキシン受容体(AhR) |
| 5.3.2. | エストロゲン受容体(ER) |
| 5.3.3. | 薬物代謝酵素 |
| 5.4. | 薬物代謝型P450分子種の課題 |
| 6. | 酸性土壌耐性植物 |
| 6.1. | 酸性土壌の成因とストレス因子 |
| 6.2. | アルミニウムストレスと耐性機構 |
| 6.2.1. | アルミニウムストレスの概要 |
| 6.2.2. | Alの毒性機構と耐性機構 |
| 6.2.3. | 低リン酸耐性 |
| 6.3. | 有機酸放出機構と有機酸放出能力強化の分子育種戦略 |
| 6.3.1. | 有機酸放出能力とAlストレス耐性 |
| 6.3.2. | 有機酸放出における有機酸代謝と有機酸細胞膜輸送 |
| 6.3.3. | 代謝工学によるクエン酸放出型酸性土壌耐性植物作出戦略 |
| 6.4. | その他の分子育種ターゲット形質 |
| 6.5. | おわりに |
|
| |
 |
植物代謝工学の工業生産への応用 |
|
| 1. | 糖質の改良と増産 |
| 1.1. | はじめに |
| 1.2. | デンプンの構造 |
| 1.3. | デンプンの生合成 |
| 1.4. | デンプンの改変 |
| 1.4.1. | 量を変える |
| 1.4.2. | 質を変える |
| 1.5. | おわりに |
| 2. | セルロース・ヘミセルロース |
| 2.1. | 植物多糖類生合成前駆体 |
| 2.2. | 糖ヌクレオチドの代謝と制御 |
| 2.3. | セルロースの生合成 |
| 2.4. | カロース(1,3-β-グルカン)の生合成 |
| 2.5. | キシログルカンの生合成 |
| 2.6. | キシランの生合成 |
| 2.7. | グルコマンナンの生合成 |
| 2.8. | キシログルカンエンドトランスグルコシラーゼ(XET) |
| 3. | リグニン |
| 3.1. | はじめに |
| 3.2. | リグニンの分子育種の幕開け |
| 3.3. | 遺伝子組換えによるリグニン組成の制御 |
| 3.4. | 新しいリグニン生合成経路の発見 |
| 3.5. | リグニン分子育種の最近の知見 |
| 3.6. | おわりに |
| 4. | アミノ酸 |
| 4.1. | 植物のアミノ酸生合成と輸送 |
| 4.2. | 植物によるアミノ酸の生産 |
| 4.2.1. | トリプトファン |
| 4.2.2. | リジンの生産 |
| 4.2.3. | アスパラギン酸の生産 |
| 4.2.4. | メチオニンの蓄積 |
| 4.2.5. | トレオニンの蓄積 |
| 5. | 脂肪酸 |
| 5.1. | はじめに |
| 5.2. | 高等植物における脂質生合成経路 |
| 5.2.1. | 脂肪酸合成酵素複合体による脂肪酸合成 |
| 5.2.2. | 2つの経路によるグリセロ脂質生合成 |
| 5.2.3. | 脂肪酸の不飽和化 |
| 5.2.4. | 脂肪酸の鎖長延長 |
| 5.2.5. | 貯蔵脂質の合成 |
| 5.3. | 脂肪酸の質の改変 |
| 5.3.1. | ラウリン酸生産ナタネ |
| 5.3.2. | 高ステアリン酸含有ナタネ |
| 5.3.3. | 高エルカ酸(C22:1)ナタネ |
| 5.3.4. | 高オレイン酸生産ダイズ |
| 5.3.5. | 高リシノール酸植物 |
| 5.3.6. | γ-リノレン酸生産植物 |
| 5.3.7. | タンパク質工学によるC18:0-ACP不飽和化酵素の改変 |
| 5.3.8. | 植物における超長鎖脂肪酸の生産 |
|
|
| 5.4. | 高等植物において高度不飽和脂肪酸を生産するための戦略 |
| 5.4.1. | 真菌類の高度不飽和脂肪酸生合成系 |
| 5.4.2. | 海洋性細菌やラビリンチュラ類の高度不飽和脂肪酸合成系 |
| 5.4.3. | 組換え植物による高度不飽和脂肪酸生産の実際 |
| 5.5. | 今後の展望 |
| 6. | テルペノイド |
| 6.1. | はじめに |
| 6.2. | テルペノイドの分類 |
| 6.3. | テルペノイドの生合成 |
| 6.3.1. | テルペノイド生合成の流れ |
| 6.3.2. | イソペンテニル二リン酸(IPP)の生合成 |
| 6.3.3. | テルペノイド生合成における鎖長延長反応 |
| 6.4. | おわりに |
| 7. | 二次代謝産物 |
| 7.1. | アルカロイド |
| 7.2. | カロテノイド |
| 7.2.1. | はじめに |
| 7.2.2. | イソプレノイド基本生合成経路 |
| 7.2.3. |  のカロテノイド生合成遺伝子群の単離と機能解明 のカロテノイド生合成遺伝子群の単離と機能解明 |
| 7.2.4. | 海洋細菌のアスタキサンチン生合成遺伝子群の単離と機能解明 |
| 7.2.5. | 真正細菌のカロテノイド生合成遺伝子と生合成酵素のプロフィール |
| 7.2.6. | 植物のカロテノイド生合成遺伝子と生合成経路 |
| 7.2.7. |  遺伝子の導入による植物の代謝工学 遺伝子の導入による植物の代謝工学 |
| 7.2.8. |  遺伝子の導入による植物の代謝工学 遺伝子の導入による植物の代謝工学 |
| 7.2.9. | 複数遺伝子の導入による植物の代謝工学 |
| 7.2.10. | おわりに |
| 8. | 酵素・生理活性タンパク質 |
| 8.1. | はじめに |
| 8.2. | 産業用酵素および分析用酵素の現状 |
| 8.3. | バイオ医薬品の現状と開発状況 |
| 8.4. | 遺伝子組換えタンパク質生産システムの比較 |
| 8.5. | 植物による酵素、生理活性タンパク質の生産の現状 |
| 8.6. | おわりに |
| 9. | 花き育種 |
| 9.1. | はじめに |
| 9.2. | 遺伝子組換えによる花色の改変 |
| 9.2.1. | 遺伝子組換えによる花色の改変例 |
| 9.2.2. | 遺伝子組換えによる青い花の作出 |
| 9.3. | 遺伝子組換えによる花の寿命の延長 |
| 9.4. | その他の形質の改変 |
| 9.4.1. | 花型の改変 |
| 9.4.2. | 草姿の改変 |
| 9.4.3. | 病虫害抵抗性等 |
| 9.5. | 今後の課題と展望 |
|
| |
 |
植物代謝工学の環境浄化、修復への応用 |
|
| 1. | 大気汚染物質 |
| 1.1. | 窒素酸化物 |
| 1.1.1. | なぜ植物利用か―ある試算の意味するもの |
| 1.1.2. | 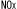 汚染の現状と環境への影響 汚染の現状と環境への影響 |
| 1.1.3. | 植物に取り込まれた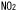 はどのように代謝されるか はどのように代謝されるか |
| 1.1.4. | 植物の同化能の多様性と遺伝子 |
| 1.1.5. | 遺伝子操作で同化能を向上させる |
| 1.1.6. | 将来の課題:ガス→ガス変換植物 |
| 1.1.7. | 植物を植えてどれくらいを減らせるか |
| 1.2. | 無機硫黄化合物 |
| 1.2.1. | 無機硫黄性大気汚染物質に対する植物の抵抗性機構 |
| 1.2.2. | 遺伝子組換え技術による有害硫黄化合物耐性の付与 |
| 2. | 重金属汚染土壌―重金属の植物に与える影響と環境修復植物の開発― |
| 2.1. | はじめに |
| 2.2. | 重金属の役割、生体への影響 |
| 2.3. | 生体内への取り込み、作用 |
| 2.4. | 金属イオンの分配、局在化 |
| 2.5. | トランスジェニック植物 |
| 2.6. | まとめ |
| 3. | アルミニウム耐性 |
| 3.1. | 酸性土壌とアルミニウム |
| 3.2. | アルミニウム毒性 |
| 3.2.1. | アルミニウム障害 |
| 3.2.2. | 根の伸長阻害を受ける部位 |
| 3.2.3. | Al集積部位と集積量 |
| 3.2.4. | アポプラスト |
| 3.2.5. | 原形質膜 |
|
|
| 3.2.6. | 細胞分裂阻害 |
| 3.2.7. | カロース |
| 3.3. | アルミニウム耐性機構 |
| 3.3.1. | アルミニウム排除機構 |
| 3.3.2. | 細胞内耐性機構 |
| 3.3.3. | アルミニウム耐性の遺伝的側面 |
| 4. | ファイトレメディエーション(植物による環境浄化) |
| 5. | 富栄養化の防止 |
| 5.1. | 窒素・リンの排出と富栄養化のメカニズム |
| 5.2. | 富栄養化対策のための植物浄化施設 |
| 5.3. | 水質浄化植物による富栄養化対策とデータベース化 |
| 5.4. | 収穫した植物の有効利用法の開発 |
| 5.5. | 浄化植物への育種の試み |
| 5.6. | 植物浄化法の課題 |
| 6. | 環境植林 |
| 6.1. | はじめに |
| 6.2. | 早成樹種による商業植林 |
| 6.2.1. | ブラジル |
| 6.2.2. | オーストラリア |
| 6.2.3. | パプアニューギニア |
| 6.2.4. | ベトナム |
| 6.2.5. | 中国 |
| 6.3. | 環境植林 |
| 6.4. | 森林の二酸化炭素(吸収能) |
| 6.5. | 地球における濃度の変化 |
| 6.6. | 森林の公益的機能 |
| 6.6.1. | 評価方法 |
| 6.6.2. | 貨幣評価しなかった森林の持つ公益的機能 |
| 6.6.3. | 算出根拠 |
| 6.7. | まとめ |
|
| |
 |
植物バイオテクノロジーの将来展望 |
|
| 7.1. | リスクアセスメント |
| 7.1.1. | はじめに |
| 7.1.2. | 安全性 |
| 7.1.3. | 生態系への影響 |
| 7.1.3.1. | 雑草化 |
| 7.1.3.2. | 遺伝子の水平移動 |
| 7.1.3.3. | 抵抗性生物の出現 |
| 7.1.3.4. | 目的外の生物への影響 |
| 7.1.4. | リスクの定量化 |
| 7.1.5. | リスクの管理 |
| 7.1.6. | 遺伝子組換え植物のリスク評価 |
| 7.1.7. | 持続可能な社会の構築 |
| 7.2. | 遺伝子組換え植物の生態系への影響 |
| 7.2.1. | はじめに |
| 7.2.2. | 懸念されている外来植物による生態系への影響 |
|
|
| 7.2.3. | 遺伝子組換え植物の生態系への影響 |
| 7.2.4. | 遺伝子組換え植物の生態系への悪影響への懸念 |
| 7.2.5. | 国際的な規制の枠組み |
| 7.2.6. | わが国の対応 |
| 7.2.7. | まとめ |
| 7.3. | 持続可能な社会 |
| 7.3.1. | 地球の変遷 |
| 7.3.2. | 地球の定員 |
| 7.3.3. | 20世紀の経済発展 |
| 7.3.4. | 食糧と水資源の危機 |
| 7.3.5. | 食糧増産の道 |
| 7.3.6. | 水資源問題の対策 |
| 7.3.7. | 植物による工業原料の生産 |
| 7.3.8. | 持続可能な社会 |
|
| |
|
|
| 1. | 遺伝子組換え植物の立場 |
| 2. | 遺伝子組換え技術を基盤とした植物代謝工学 |
|
|
| 3. | ストレス抵抗性との組合せが重要 |
| 4. | 植物代謝工学の未来 |
|
| |
|
|